|
|
|
|
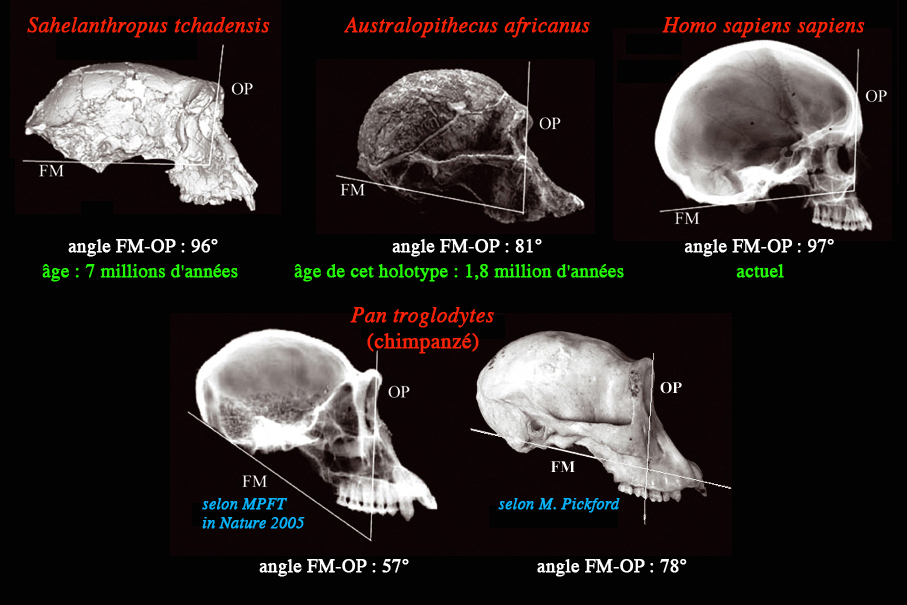
Orbital planes (OP) and foramen magnum planes (FM) of african
apes and hominids according to MPFT (Nature, 2005, La Recherche 2005, ...) and Martin Pickford (Anthropologie, 2005).
.ORIENTATION OF THE FORAMEN MAGNUM IN LATE MIOCENE TO EXTANT AFRICAN
APES AND HOMINIDS
ANTHROPOLOGIE
XLIII/23 pp. 191198 2005
Martin Pickford
ABSTRACT : The postcranial skeleton of the Late Miocene hominoid Sahelanthropus tchadensis is unknown ; because of this the possibility of bipedal posture
and locomotion has been based on cranial features, an interpretation
that has not been accepted by all paleoanthropo-logists. Part
of the evidence includes the orientation of the foramen magnum
relative to the orbital plane (FM-OP angle), an acute angle being
said to characterise more pronograde postures such as that of
the chimpanzee, and more obtuse angles to characterise orthograde
humans, australopithecines and Sahelanthropus. This paper examines the soundness of the FM-OP evidence used
to infer the likelihood of bipedal posture in Sahelanthropus.
KEY WORDS : Hominoids Foramen magnum plane (FM) Orbital plane (OP)
FM-OP angle Occlusal plane Miocene to recent Bipedal posture
INTRODUCTION
Zollikofer et al. (2005: 758, fig. 4) employed the relationship
between the plane of the foramen magnum and the orbital plane
(FM-OP angle) to infer that "as a quadruped" Sahelanthropus "would require an unusually extended angle of the neck relative
to the plane of the foramen magnum", an inference reinforced by
Brunet and Allemand (2005: 37) who, on the basis of the FM-OP
angle, wrote that "the probability that Sahelanthropus tchadensis was a biped is greater than that it was not" (present author's
translation from the French).
Zollikofer et al. (2005) reported that FM-OP angle in Pan troglodytes is 63.7°+/6.2°, n=20, and in humans 103.2°+/6.9°, n=23. In
their figure 4, the authors illustrated the two planes superimposed
on the skulls of a) a human, b) a chimpanzee, c) Australopithecus africanus (Sts 5) and d) Sahelanthropus tchadensis. Brunet and Allemand (2005: p. 38) published the same figure
and reported that the chimpanzee FM-OP "is acute, of the order
of 65°".
Measurement of the FM-OP angle in the figure reveals that the
human one is 97°, that of the chimpanzee is 57°, that of the australopithecine,
81° and that of Sahelanthropus, 96°. Out of the four species illustrated, the FM line intercepts
the dentition in two, and only in the chimpanzee is the FM line
below the dentition. The OP in three of the species intercepts
the molar row (in chimpanzee the intercept is M1/, in Sahelanthropus it is the junction between M1/M2/, and in the australopithecine
it is about M2/), humans being the exception with the OP intercepting
the premolar row at P3/.
The human FM-OP angle illustrated (97°) is close to the lowest
value measured in humans (96.3° according to Zollikofer et al.
2005) and the chimpanzee FM-OP illustrated (57°) is more acute
than the most acute angle given in the text (57.5°) and well below
the mean (63.7°) given by these authors or the value (ca 65°)
estimated by Brunet and Allemand (2005). Because of the latter
inconsistency, the present author decided to measure the FM-OP
angle in chimpanzees, gorillas and some humans and to compare
them with the values published for Sahelanthropus and other hominoids. This paper presents the results of this
analysis and discusses the implications for bipedal posture in
Sahelanthropus.
MATERIAL AND METHODS
The skulls of 21 chimpanzees (10 male, 11 female), 30 gorillas
(19 male, 11 female) and 2 humans (1 unrecorded sex, 1 female)
(Table 1) were photographed using a digital camera in lateral
view with a straight edge positioned in contact with opisthion
and basion (Figure 1). The resultant images were treated using
Photoshop to eliminate unwanted elements of the image such as
the background. A line was drawn precisely along the edge of the
ruler that was in contact with opisthion and basion, and the image
of the ruler was then eliminated (Figures 2, 3, 4). This procedure
gives an accurate estimate of the FM plane (+/ 0.25°), opisthion
and basion being precise cranial landmarks located in the sagittal
plane.
The OP is inherently a less accurate plane to measure on account
of the lack of precise landmarks. Zollikofer et al. (2005) and
Brunet and Allemand (2005) did not provide details of how they
measured the orientation of the OP, but judging from the illustrations
it was obtained by drawing a vertical line in the middle of the
orbit extending from the anterodorsal margin of the orbit to the
antero-ventral edge of the orbit immediately beneath the former
point. There are four main difficulties with this measurement,
the first being that the left and right sides of the same skull
may be different (in fact each orbit has its own OP). The second,
and a more serious difficulty is that the upper and lower margins
of the orbits are curved dorsoventrally (strongly) and mediolaterally
(gently), and thus it is difficult to define precise points for
consistent measurements between individuals or successive measurements
in the same individual, and thus of obtaining reliable results
for the orientation of the orbital plane itself. I estimate that
the uncertainty in defining the margins of the orbit results in
an error margin of 35°. The third difficulty is that the orbits
are not in the sagittal plane, and thus the FM line and the OP
lines are offset from each other, intersecting each other outside
the points used to define the OP. The fourth difficulty relates
to the images of the skulls obtained, a slight difference in position
of the camera introducing a small difference in apparent orientation
of the OP. To minimise the effect, images were taken such that
the orbits were in the same plane as each other and the camera
was positioned so as to be near the intersection of the FM and
OP planes (error margin in OP estimated to be +/ 1°). Thus, with
the method used, the FM plane can be accurately defined (error
margin +/ 0.25°) whereas the OP less so (error margin +/ 46°).
The OP was drawn onto the digital images so obtained, and the
angle between the FM-OP measured using a protractor.
The FM-OP angles published by Zollikofer et al. (2005) of chimpanzees
(63.7° +/ 6.2°) and humans (103.2° +/ 6.9°) are markedly divergent,
with no overlap even if the maximal error margins are applied.
The error margins introduce a relatively minor degree of uncertainty
(noise) into the analysis but do not completely invalidate the
approach.
RESULTS
Chimpanzee foramen magnum
In the chimpanzee skulls examined (Table 1) the FM line intersects
the occlusal plane in 20 individuals, and in only one individual
(1952.144, male) does it lie below the tooth row (i.e. the FM
intersects the occlusal plane behind the M3/). In chimpanzees
the orientation of the FM line is extremely variable, in some
individuals the line extends anteriorly to the level of the incisors,
in others it intersects the molar row (Figure 2).
Chimpanzee orbital plane
In chimpanzees the orbital plane (OP) intercepts the M1/ in 16
individuals, the M2/ in 4 individu-als, and in only one individual
(1952.144, male) does it intercept the M3/.
Chimpanzee FM-OP angle
The most acute chimpanzee FM-OP angle measured was 60°, the least
acute 78°, mean 69°. Males (60°76°) tend to have a more acute
FM-OP than females (64°78°) but there is a great deal of overlap
between the sexes in the sample analysed.
Gorilla foramen magnum
As with the chimpanzee skulls, in most gorillas examined (Table
1), the FM line intercepts the occlusal plane, with only four
individuals (all males) having the line below the tooth row (ie
intersecting the occlusal plane behind the M3/). In some females
the FM line is obtuse enough to intercept the incisors (Figure
3).
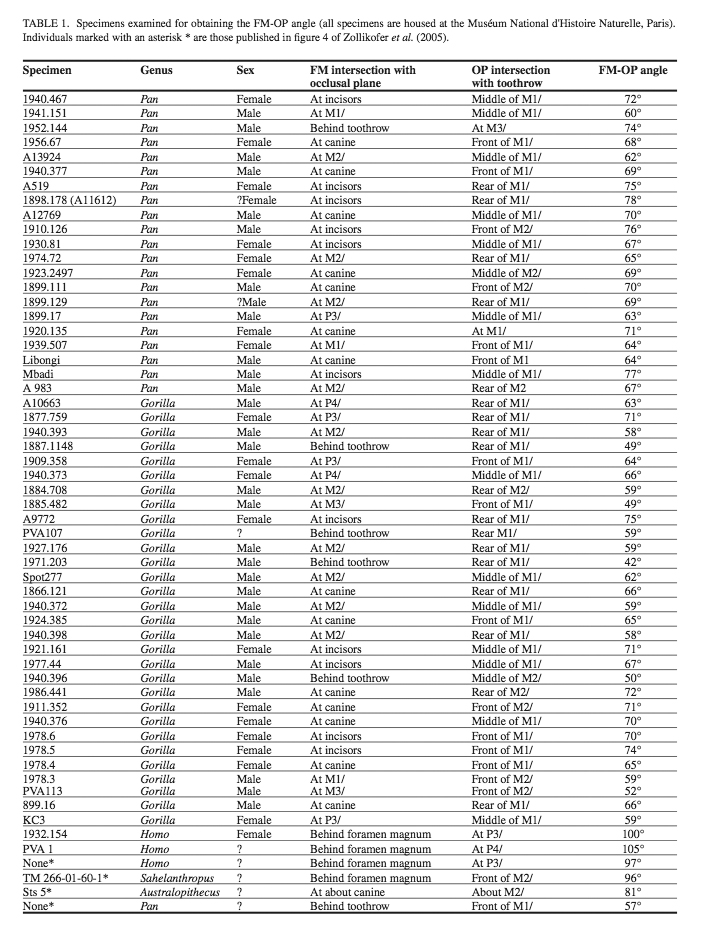
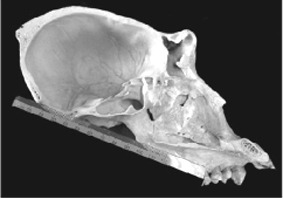
FIGURE 1. Method of determining the orientation of the foramen
magnum plane (FM). A straight edge is laid in contact with basion
and opisthion, this line defining the FM plane which is at right
angles to the sagittal plane. Specimen KC3 female Gorilla.
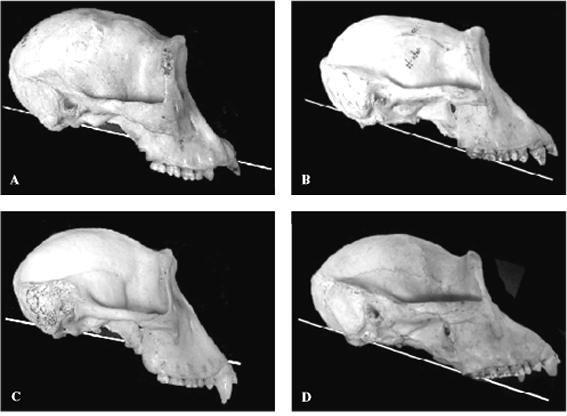
FIGURE 2. Examples of the FM plane in extant chimpanzees showing
part of the range of variation. Note that in most specimens the
FM plane intersects the toothrow. A 1898.178 female; B 1974.72,
female; C 1910.126 male; D A 13924 male.
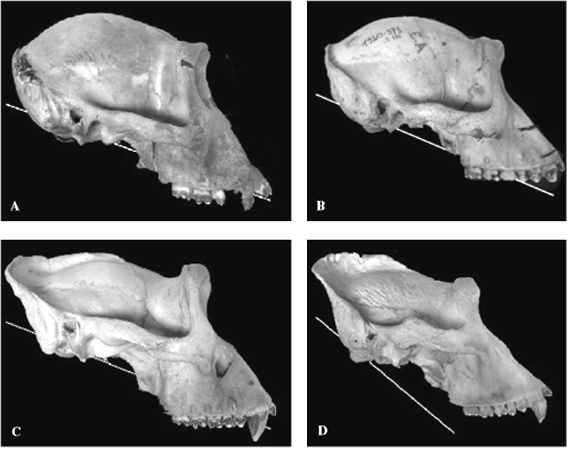
FIGURE 3. Examples of the FM plane in extant gorillas showing
part of the range of variation. Males tend to have steeper FM
planes than females, but note that in most individuals the FM
plane intersects the tooth row. A 1978.6 female; B 1910.373
female; C 1924.385 male; D 1971.203 male.
Gorilla orbital plane
The orbital plane intercepts the M1/ in the majority of gorilla
individuals (23) or is anterior to it (at the front of M1/ in
7 females). It intersects M2/ in 6 individuals. Thus the orbital
plane of the gorilla has much the same relation to the tooth row
as it does in chimpanzees.
Gorilla FM-OP angle
The most acute gorilla FM-OP angle measured was 42° (2 males)
and the least acute was 75° (a female), the mean being 63.5°.
Males (42°72°, mean 57°) tend to have more acute FM-OP angles
than females (59°75°, mean 67°) but there is overlap between
the sexes. Aged males have more acute FM-OP angles than young
males, and it is evident that this angle changes ontogenetically.
Human foramen magnum
In both of the humans measured (Table 1) the FM line was well
above the dentition, intersecting the nasal cavity. This is considerably
more elevated anteriorly than in any of the chimpanzees and gorillas.
The relationship with the occlusal plane is thus diametrically
opposite to that of chimpanzees and gorillas (Figure 4), the intersection
of the FM plane occurring posterior to the foramen magnum rather
than anterior to it as in chimpanzees and gorillas.
Human orbital plane
The orbital plane in both humans examined intersected the premolars.
Human FM-OP angle the two humans in the sample have FM-OP angles of 100°
and 105°.
COMPARISONS
Foramen magnum orientation
In chimpanzees and gorillas, the plane of the foramen magnum,
as defined by basion and opisthion, generally intersects the occlusal
plane somewhere along the tooth row, the few specimens that intersect
this plane behind the M3/ all being males (1 chimpanzee out of
20 examined, and 4 out of 31 gorillas included in the study).
The specimens in which the FM plane falls below the toothrow are
all fully adult to aged males, young males being more like females
in the values obtained. From this it is inferred that the relationship
between the FM plane and the maxilla changes ontogenetically,
the inclination of the FM increasing with age relative to the
occlusal plane, especially in gorillas (Delattre, Fenart 1960).
It is also evident that there is a great deal of individual variation
in the orientation of this plane, reflecting individual differences
in the angle at which the splanchnocranium is hafted onto the
neurocranium. In humans the FM plane in the two specimens examined
lies above the toothrow (including the roots), intersecting the
nasal cavity anteriorly and the occlusal plane well behind the
foramen magnum (Figure 4). In the individual illustrated by Zollikofer
et al. (2005) the FM intersects the apices of the molar roots,
and thus appears to be an extreme variant among humans, a conclusion
confirmed by the values published by the authors.
The FM plane in Sahelanthropus as reconstructed by Zollikofer et al. (2005), lies above the
roots of the toothrow including the apex of the canine, which
is extremely elongated within a hominid context, and it intersects
the nasal cavity anteriorly and the occlusal plane well behind
the foramen magnum as in humans.
The FM plane of the australopithecine illustrated by Zollikofer
et al. (2005) is inclined at such an angle that it intersects
the tooth row low down, much lower than in the Sahelanthropus reconstruction and close to the condition in Gorilla and Pan.
The published FM plane of Sahelanthropus relative to the occlusal plane is more elevated than it is in
australopithecines, chimpanzees and gorillas, and falls well within
the range of variation of humans.
The orbital plane
The orbital plane of chimpanzees and gorillas generally intersects
the M1/, with a few individuals intersecting the M2/ behind or
the contact between M1/ and P4/ anteriorly. In none of the chimpanzees
examined did the OP intersect the premolars, although in some
female gorillas the plane intersected the contact between the
P4/ and M1/.
In humans, the OP intersects the premolars, usually the P3/ (Zollikofer
et al. 2005) (Figure 4).
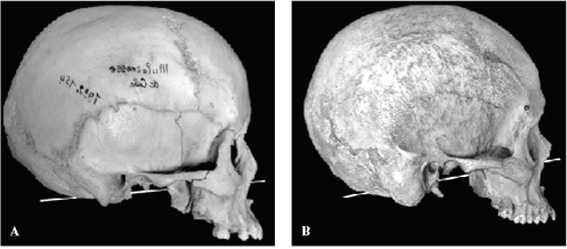
FIGURE 4. Examples of the FM plane in humans. Note that the plane
does not intersect the toothrow. A 1932.152 female (reversed);
B PVA 1, unrecorded sex.
In the reconstruction of Sahelanthropus, the OP intersects the M2/, which is posterior to the mean position
in chimpanzees and gorillas, although it falls within the range
of variation of these species. Thus the intersection of the OP
and the toothrow of Sahelanthropus is markedly different from the situation in humans.
In Australopithecus, the OP published by Zollikofer et al. (2005) intersects the
toothrow far back, probably about the level of M2/ (the teeth
are missing in Sts 5).
Thus, if the OP of Sahelanthropus is correctly positioned in the reconstruction, then its orientation
is more like that of chimpanzees and gorillas than that of humans.
FM-OP angle
The FM-OP angle reflects the position and orientation of two planes.
In the reconstructed cranium of Sahelanthropus, the FM plane falls well within the range of variation of humans,
whereas the OP falls into the lower part of the range of variation
of gorillas and chimpanzees, well outside the range of variation
of humans. As a consequence, the angle between these planes, as
published by Zollikofer et al. (2005), falls close to the human
range of variation, and well above that of gorillas, chimps and
australopithecines.
In Australopithecus, both the FM and OP published by Zollikofer et al. (2005) fall
within the range of variation of gorillas and chimpanzees, although
the angle between them is slightly outside the range of variation
of the African apes.
The chimpanzee FM-OP angle (57°) illustrated by Zollikofer et
al. (2005) and Brunet and Allemand (2005) is more acute than any
of the specimens examined in this study, and is smaller than the
most acute angle (57.5°) listed by Zollikofer et al. (2005) in
their text. It is 8° more acute than the value published by Brunet
and Allemand (2005).
The chimpanzee FM-OP measurements in this study range from 60°
78° (mean 69°) which is uniformly less acute than the range
of variation published by Zollikofer et al. (2005 63.7 +/ 6.2°).
These differences are possibly related to ways of measuring the
OP, taking into account the estimated error margin of 46°. The
chimpanzee individual illustrated by these authors falls beyond
their own estimate of the range of variation of the FM-OP angle
in chimpanzees, a result confirmed by this study. In particular,
the orientation of the FM plane in the illustration appears to
be too steeply inclined anteriorly, the line representing the
plane passing well below the tooth row. Out of the 20 chimpanzees
examined during the present study, only one individual showed
an FM line below the tooth row, all the others intersecting the
occlusal plane, some as high as the incisors. It is thus concluded
that the chimpanzee FM-OP angle illustrated by Zollikofer et al.
(2005) exaggerates the acuteness of the FM-OP angle and thus magnifies
the apparent difference between chimpanzees on the one hand and
the other hominoids illustrated by them on the other. Examination
of the image suggests the possibility that basion has been misidentified.
The FM-OP angle of Australopithecus africanus (Sts 5) measured directly from the illustration published by
Zollikofer et al. (2005) is 81°. This is only 3° more obtuse than
the highest value for the chimpanzee examined in this study and
11.1° greater than the most obtuse FM-OP measurement that Zollikofer
et al. (2005) provided in the text (69.9°). In Australopithecus the FM plane intersects the toothrow in much the same way as
in chimpanzees and gorillas, and it is likely that the range of
variation of the FM-OP in australopithecines is about the same
order of magnitude as it is in extant African apes. It is not
known precisely what the range of variation in australopithecines
is, but the chances are that Sts 5 has neither the most acute
nor the most obtuse FM-OP angle among the subfamily.
It is thus possible that there is overlap between the FM- OP angle
of chimpanzees and australopithecines. It is also possible that
there is overlap between the FM-OP angle of female gorillas and
australopithecines, the most obtuse gorilla FM-OP angle determined
in this study (75°) being only 6° less than the estimated angle
in A. africanus.
DISCUSSION AND CONCLUSIONS
It is generally agreed that australopithecines were bipedal, whereas
chimpanzees and gorillas are quadrupedal, albeit with the ability
to stand and move on two feet for short periods and short distances.
Australopithecines show skeletal morphology that reflects their
bipedal stance and locomotion, whereas chimpanzees and gorillas
do not. The fact that the FM-OP angles of chimpanzees and gorillas
are close to those of australopithecines, with the possibility
of overlap between the ranges of variation, indicates that this
angle does not provide a clear idea of locomotor or postural potential,
at least for these groups.
Humans in contrast possess a considerably more obtuse FM-OP than
either australopithecines or African apes (103.2 +/ 6.9° according
to Zollikofer et al. 2005). There is no overlap between the ranges
of variation of chimpanzees and gorillas on the one hand (maximum
78°) and humans (minimum 96.3°) on the other. The FM-OP value
for Sahelanthropus is estimated to be 96° by Zollikofer et al. (2005), which, if
correctly determined, would imply that it is just below the range
of variation of humans.
Given the observation that the FM plane of the reconstructed Sahelanthropus falls within the range of variation of humans, whereas its OP
falls well outside the human range, it is pertinent to ask whether
the value of the FM-OP angle illustrated by Zollikofer et al.
(2005) is reliable. The proximity of values between the FM-OP
of humans and Sahelanthropus has been used to infer the strong possibility of bipedal posture
in the genus (Brunet, Allemand 2005). If the angle is correctly
determined, and if the FM-OP angle reflects posture and locomotor
repertoire as claimed, then it is difficult to evade the conclusion
that Sahelanthropus was likely to have been better adapted for bipedal posture than
australopithecines were, as the FM- OP of Australopi-thecus is significantly more acute (81°) than that of Sahelanthropus (96°), and is only marginally more obtuse than that of chimpanzees
(most obtuse angle is 78°). However, since the OP of Sahelanthropus is quite unlike that of humans, it is legitimate to ask whether
its FM plane has been accurately reconstructed.
The original specimen of Sahelanthropus tchadensis is crushed and warped (Brunet et al. 2002), making any direct
measurement of the foramen magnum and orbital planes, as well
as the relationship between them, impossible. For this reason,
Zollikofer et al. (2005) based their measurements on a virtual
reconstruction of the skull. The basicranium was particularly
badly damaged, with the foramen magnum fractured into two, one
part with basion, the other with opisthion (Brunet et al. 2002).
Furthermore, the relationships between the basicranium on the
one hand and the rest of the neurocranium and splanchnocranium
on the other were heavily distorted, with displacement of fragments
in three dimensions, as well as rotation and translation of the
various pieces relative to each other. There could even be some
plastic deformation, although this is not immediately apparent
from the published images. Given that the basicranium is far removed
from the splanchnocranium (taking into account size differences,
the distance between the foramen magnum and the rear of the tooth
rows is as great in Sahelanthropus as it is in gorillas), then the possibility of obtaining an erroneous
relationship between the foramen magnum and the orbits is rather
high.
With crania oriented in the Frankfurt horizonal, the dorsal profile
of the skulls of chimpanzees, gorillas, australopithecines and
humans show a rise in the profile of the braincase behind the
orbits. In the virtual reconstruction of Sahelanthropus this part of the cranium is almost flat, suggesting that the
neurocranium may be hafted onto the splanchnocranium at too horizontal
an angle. If this is so, then all the posterior elements of the
neurocranium may have been rotated ventrally with respect to their
life position (i.e. in right lateral view, counterclockwise rotation
of the neurocranium relative to the splanchnocranium). There are
several breaks in the neurocra-nium about which rotation could
have taken place. Removal of this rotation would make the nuchal
area steeper and would make the plane of the foramen magnum more
acute relative to the orbital plane. A change in angle of the
FM plane of only 5° would result in a value of the FM-OP angle
of ca 89°, a change of 10° would result in an angle of ca 84°,
not very different from the maximal figure for chimpanzees and
gorillas. Given the uncertainties of working with crushed and
distorted fossils, and considering the ape-like orientation and
position of the orbital plane in Sahelanthropus, the presence
of a fully human-like FM plane in it is surprising.
Because, if Sahelanthropus is a hominid which gave rise to humans via australopithecines,
as envisaged by Brunet et al. (2002), Zollikofer et al. (2005)
and Brunet and Allemand (2005), then the FM plane would change
from human-like in Sahelanthropus tchadensis to ape-like in Australopithecus and back to human-like in the genus Homo, at the same time that its OP remained ape-like until the human
stage was reached.
In view of these contradictions it is considered that the FM plane
of Sahelanthropus has been incorrectly estimated by Zollikofer et al. (2005). In
the author's view, the FM of Sahelanthropus would probably have intersected the toothrow as in chimpanzees,
gorillas and australopithecines, and if so, then its FM-OP angle
would have been appreciably more acute than the reconstructed
figure published by Zollikofer et al. (2005). As a consequence
it is considered premature to deduce bipedal posture and locomotion
for Sahelanthropus on the basis of its reconstructed FM-OP angle. This conclusion
reinforces that of Wolpoff et al. (2002) that Sahelanthropus tchadensis was not a hominid in the strict sense of the term.
ACKNOWLEDGEMENTS
I thank Y. Coppens, B. Senut, A. Beauvilain, E. Gitonga, C. de
Muizon and S. Sen for support. Thanks to C. Lefevre for access
to skulls of African apes in her care.
REFERENCES
BRUNET M., ALLEMAND L., 2005 : Un nouveau grade d'évolution humaine.
La Recherche 387: 3643.
BRUNET M., GUY F., PILBEAM D., TAISSO MACKAYE H., LIKIUS A., AHOUNTA
D., BEAUVILAIN A., BLONDEL C., BOCHERENS H., BOISSERIE J-R., DE
BONIS L., COPPENS Y., DEJAX J., DENYS C., DURINGER P., EISENMANN
V., FANONE G., FRONTY P., GERAADS D., LEHMANN T., LIHOREAU F.,
LOUCHART A., MAHAMAT A., MERCERON G., MOUCHELIN G., OTERO O.,
PELAEZ-CAMPOMANES P., PONCE DE LÉON M., RAGE J-C., SAPANET M.,
SCHUSTER M., SUDRE J., TASSY P., VALENTIN X., VIGNAUD P., VIRIOT
L., ZAZZO A., ZOLLIKOFER C., 2002 : A new hominid from the Upper
Miocene of Chad, Central Africa. Nature 418 : 145151.
DELATTRE A., FENART R., 1960 : L'Hominisation du crâne. Editions
CNRS, Paris VII. 418 pp.
WOLPOFF M., SENUT B., PICKFORD M., HAWKS J., 2002 : Palaeoanthropology
(communication arising): Sahelanthropus or Sahelpithecus? Nature 419 : 581582.
ZOLLIKOFER C., PONCE DE LÉON M., LIEBERMAN D., GUY F., PILBEAM
D., LIKIUS A., MACKAYE H., VIGNAUD P., BRUNET M., 2005 : Virtual
reconstruction of Sahelanthro-pus tchadensis. Nature 434: 755759. |
|
