|
|
|
|
|
South African Journal of Science 100, March/April 2004
South African Journal of Science 100, September/October 2004
South African Journal of Science 101, March/April 2005
Dossier de presse :
Ann Gibbons, "The First Human. The race to discover our earliest
ancestors". (Elle omet le South African Journal of Science 101, March/April
2005)
|
South African Journal of Science 100, March/April 2004
|
La recherche en action
Nouveaux détails concernant des fossiles attribués à Sahelanthropus tchadensis (Toumaï).
Alain Beauvilain and Yves Le Guellec
Les fossiles dâge Miocène attribués à Sahelanthropus tchadensis sont dune grande importance scientifique et culturelle. Ils
sont lobjet dun débat sur leur nature dhominidé ou de grand
singe. Il est donc nécessaire de donner et de préciser les détails
de leur découverte. Nous démontrons que la troisième molaire de
la demi-mandibule droite illustrant larticle de Nature est en réalité une troisième molaire gauche et quune incisive
collectée sur le site ne figure pas dans cet article. Ces particularités
affectent le nombre minimal dindividus présents et les associations
entre les différents spécimens.
Introduction.
La découverte et lannonce de fossiles attribués à St a confirmé limportance du Miocène en Afrique comme une période
dintérêt majeur pour létude de lorigine des hominidés. Linterprétation
de Sahelanthropus comme un premier hominidé (possiblement bipède) est lobjet dun
débat dans lequel quelques auteurs déclarent quil sagit probablement
dun grand singe quadrupède. Brunet a maintenu son point de vue
que Sahelanthropus était un hominidé. En raison de la nature de ce débat et de ses
implications pour la paléoécologie et pour les origines de lhomme,
il est nécessaire de préciser les particularités regardant lhypodigme
et lanatomie de Sahelanthropus. Par conséquent, lobjectif de cette contribution est de consigner
les détails géomorphologiques du site de TM 266 et de ses environs,
qui a livré les fossiles de Sahelanthropus, ainsi que de décrire les circonstances entourant les découvertes
elles-mêmes, puisquil semble quil y ait eu quelques confusions
au sujet des associations de fossiles et même au sujet du nombre
total de spécimens mis au jour sur le site. Etant donné limportance
de cette découverte en termes dhéritage humain et de son importance
scientifique, nous jugeons nécessaire de fournir des informations
sur le contexte de ces découvertes ainsi que des détails sur les
fossiles eux-mêmes.
Le premier auteur a conduit la mission de terrain qui a abouti
à la découverte du crâne, bien connu sous le nom de Toumaï, tout
comme il a conduit les expéditions suivantes qui ont découvert
les autres fragments de mandibules et de dents isolées attribuées
à des Sahelanthropus. Il était donc intimement concerné par le déroulement des évènements.
Le contexte géologique.
Toros-Menalla est le nom donné par les paléontologues à une vaste
région de 150 km de long dans louest de lerg du Djourab au Tchad.
Lun des faits remarquables de la partie centrale de cette zone
est lexistence dun talus de faible dénivelé, généralement dune
dizaine de mètres, et de 40 km de long (figure 1). Cet escarpement,
faisant face à lEst, est nettement indiqué sur la carte topographique
IGN NE-33-VI au 1/200.000 sur une distance de plus de 25 km (de
16° 20 N ; 17° 15 E à 16° 15 N ; 17 28 E). Ce relief représente
peut-être une ancienne faille réactivée par les forces tectoniques
liées au volcanisme du Tibesti. LEmi Koussi (un cône volcanique
de 80 km de diamètre culminant à 3415 m, formé depuis la fin du
Tertiaire, du Miocène, jusquà une époque sub-contemporaine) est
seulement à 400 km. Aujourdhui, des témoignages dactivité tectonique
entourent le Djourab comme sur les marges nord du bassin tchadien
où la falaise de lAngamma atteste dune flexure profonde.
Il est regrettable que la zone ait été décrite comme une plaine
désolée dont la monotonie est seulement interrompue par les dunes
modernes (4) qui donne une impression erronée du potentiel de
la zone pour la stratigraphie et la paléontologie. Une semblable
impression restrictive est donnée par la coupe stratigraphique
publiée par Vignaud et al (5), qui montre un épaisseur de six mètres de sédiments constitués
de seulement deux unités stratigraphiques. Il faut indiquer que
cette coupe a été réalisée à TM 266 mais que, sur les 40 km du
talus, la succession des strates est réellement plus épaisse et
plus complexe que cela avec des dépôts lacustres, péri-lacustres,
sub-aériens, fluviaux et éoliens (figure 1).
Le talus forme soit un escarpement unique, soit une succession
de petites terrasses, telles des marches, quelquefois séparées
lune de lautre par plusieurs centaines de mètres. Même sil
est nivelé à ses deux extrémités, dans sa partie centrale la combinaison
de la corrasion et de la déflation par le vent a, par endroits,
dégagé un beau talus ou une série de replats et de rejets structuraux
(figure 1). Là, la partie sommitale est riche de preuves géomorphologiques
et archéologiques indiquant les rivages de la dernière phase du
méga lac Tchad. Toutefois, les sédiments de la base du talus sont
très vieux, datés de la fin du Miocène sur la base de létude
biochronologique des fossiles de mammifères, et non apparentés
au Pléistocène et à lhistoire récente du lac Tchad mais plutôt
à un prédécesseur bien plus vieux.
Pour donner une idée de la richesse paléontologique de la zone,
il suffit de remarquer que de la première reconnaissance du talus
en juillet 2001 à mars 2002, 59 sites fossilifères ont été cartographiés
qui ont livré 1627 fossiles. Ils représentent 20% de lensemble
des fossiles catalogués depuis 1994 par le projet scientifique.
Parmi ces fossiles, outre Sahelanthropus tchadensis, de nouveaux taxons de Giraffidæ, Hippopotamidæ et Anthracotheriidæ ont été mis au jour. La collecte porte également sur une abondante
microfaune et dautres mammifères, aussi bien aquatiques que terrestres,
tels que des Deinotherium, des Anancus et des poissons. Parmi ces sites, cest TM 266 qui, avec 691
fossiles (à mars 2002) qui compte la faune la plus variée. Le
potentiel fossilifère de cette zone demeure important, notamment
pour la mise au jour de petits fossiles mais aussi en raison de
lextension de la surface couverte par une fine épaisseur de sable
ou par les dunes que le vent peut déplacer.

Fig. 1. Vue de la face est du talus de Toro-Menalla (site TM 039),
dans lerg du Djourab au Tchad, montrant les dépôts dâge Miocène
affleurant en de petits escarpements (habituellement composés
de grès et dautres couches résistantes telles que des diatomites),
recouverts par les sables des dunes actuelles. Lérosion dans
cette zone est largement due à la déflation éolienne mais il est
considéré que le talus doit son origine à la présence dune ligne
de faille, qui est cachée sous les sables en contrebas du talus.
Notons que lépaisseur totale des terrains du Miocène de cette
région excède grandement les dix mètres. En dépit de la possibilité
de remaniement des fossiles, les faunes mammaliennes collectées
comprennent exclusivement des taxons de la fin du Miocène.
Paléoanthropologie.
Hypodigme de Sahelanthropus tchadensis.
Il existe quelques différences entre la liste des fossiles attribués
à Sahelanthropus tchadensis dans larticle de Nature (1) et celle que nous avons publiée sur internet (6) à partir
des spécimens collectés par le premier auteur, ses collaborateurs
tchadiens et Laurent Viriot, elle-même établie à partir de linventaire
de terrain établi au moment des découvertes (7). Le compte-rendu
qui suit traite des fossiles dans lordre chronologique de leur
découverte. Leur catalogage comprend quatre sections : la première,
TM 266, fait référence au site ; la seconde indique lannée de
découverte ; la troisième est leur position dans lordre de leur
découverte ; et la quatrième (si elle existe) dénombre les différents
éléments dun même fossile.
TM 266-01-060-1 (le crâne) a été mis au jour simultanément avec
TM 266-01-060-2 (un fragment de symphyse avec les alvéoles des
incisives et des canines) le 19 juillet 2001 par Ahounta Djimdoumalbaye.
Provenant du même endroit, une incisive non publiée a été trouvée
le 21 décembre 2001 à NDjaména par Laurent Viriot lors du tri
des résidus de tamis. Ce spécimen est une demi-incisive, coupée
verticalement du sommet de la couronne à la base de la racine
(figure 2 document 4). Cette dent diffère de lincisive supérieure
(TM 266-01-448) dont la photographie a été publiée dans Nature (référence 1, figure 2.a).
Il convient de signaler que, depuis la découverte de la mandibule
dAustralopithecus bahrelghazali (8) et par respect pour ces témoins de lhistoire de nos origines,
il est une pratique de la Mission Paléoanthropologique Franco-Tchadienne
de ne pas écrire sur les spécimens attribués à des hominidés fossiles
mais sur linventaire de terrain. Le catalogage des spécimens
TM 266-01-447 (une molaire dans la référence 1) et 448 apparaît
pour la première fois dans la base de données des fossiles établie
sur CD par luniversité de Poitiers et pour la seconde fois dans
larticle de Nature de juillet 2002. Nous ignorons le spécimen TM 266-01-447 et les
sites et circonstances exactes de la découverte des fossiles TM
266-01-447 et 448.
Le 1er novembre 2001, la canine supérieure droite complète de
TM 266-01-60-1 était trouvée, lors du tamisage de lemplacement
exact du crâne, par Fanoné Gongdibé. La dent consiste en deux
fragments qui simbriquent parfaitement sur la racine de la canine
droite.
Les fossiles TM 266-02-154-1 (une demi-mandibule droite portant
P4 à M2 et les racines de P3 et M3) et TM 266-02-154-3 (couronne dune M3 gauche) ont été découverts le 20 janvier 2002, à environ 105
mètres au nord-nord-est du site du crâne de Toumaï. Cette mandibule
est lobjet de deux photographies dans larticle de Nature (référence 1, figures 2b et c), lune en vue occlusale, lautre
sous forme dune coupe axiale de scanner.
Le spécimen TM 266-02-154-2 (canine inférieure ; référence 1,
figures 2d et e) a été découvert le 30 janvier 2002 à NDjaména
lors du tri des résidus de tamisage de lendroit où ont été trouvées
la mandibule et la M3. Il était complètement encroûté avec du sable et ne pouvait donc
être photographié au moment de sa découverte. Enfin, le spécimen
TM 266-02-203 (une symphyse mandibulaire avec prémolaire, racines
de canine et alvéole dincisive) était mis au jour le 12 mars
2002 à une douzaine de mètres de la mandibule et de la M3.
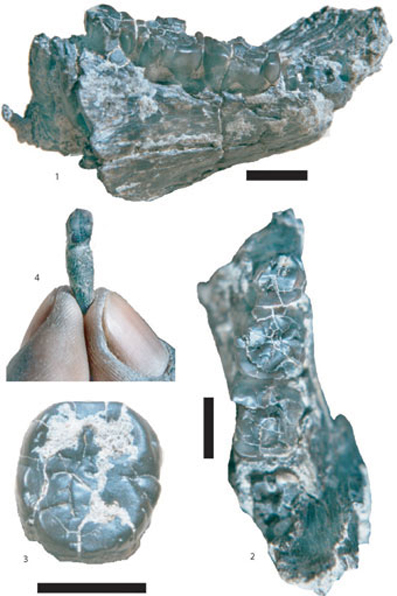
Fig. 2. Fossiles attribués à Sahelanthropus tchadensis, issus du site TM 266, Toros-Menalla, Tchad. Barre déchelle
: 10 mm.
(1) TM 266-02-154-1, vue oblique côté lingual de l hémi-mandibule
droite montrant le spécimen au moment de sa découverte, sans la
couronne de la m/3, et montrant les racines de la P3.
(2) TM 266-02-154-1, vue occlusale de lhémi-mandibule droite
au moment de sa découverte montrant les racines de la M3.
(3) TM266-02-154-3, vue occlusale de la M3 gauche au moment de sa découverte et avant que le spécimen soit
nettoyé (partie mésiale en haut).
(4) TM 266-01-448 ?, incisive endommagée non publiée trouvée à
proximité du crâne holotype de Sahelanthropus tchadensis.
Anatomie.
La photographie publiée dans Nature de lhémi-mandibule droite TM 266-02-154-1 (référence 1, figure
2b, c) montre la rangée dentaire complète de P4 à M3. En vue occlusale, la M3 apparaît comme une dent bien préservée alors que les trois autres
dents sont très endommagées, éclatées. A linverse, sur la tomographie,
ce sont les racines de cette M3 qui apparaissent abîmées, les deux racines étant cassées. En
fait, la M3, dont seule la couronne est préservée, a été trouvée isolément
à quelques décimètres de lhémi-mandibule. Elle a été cataloguée
postérieurement avec le numéro dordre 3 par analogie avec les
pièces 1 (la mandibule) et 2 (la canine), les autres pièces attribuées
à TM 266-02-154, unique référence de linventaire de terrain.
Cette M3 a été fixée ultérieurement sur lhémi-mandibule au laboratoire
de luniversité de Poitiers.
La figure 2 (1 et 2) montre la mandibule telle quelle était le
jour de sa découverte. Notons que la troisième molaire de droite
est manquante. Les racines de la P3 sont clairement visibles sur la partie antérieure de la vue oblique
(figure 2 (1)), senfonçant profondément dans le corps de la mandibule
en sincurvant vers larrière. Lextrême position buccale de la
racine mésiale (à gauche sur la figure) indique que la racine
de la canine était probablement de grande taille et que son extrémité
était positionnée de manière appréciable, plus à larrière que
son collet. La grande taille de la racine de la canine indique
que cette mandibule appartenait vraisemblablement à un mâle.
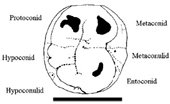
Fig. 3. Interpretation de la M3 gauche TM 266-02-154-3. La disposition des protoconide, hypoconide,
hypoconulide, metaconide et entoconide montre sans équivoque que
cette molaire inférieure est une gauche. Barre déchelle : 10
mm.
La vue occlusale de la dent TM 266-02-154-3 évoque immédiatement
une molaire inférieure dhominoïde (figure 2 (3)). Les cinq cuspides
et le diamètre mésio-distal de la face occlusale plus grand que
le diamètre bucco-lingual et la face mésiale (en haut de la figure
2 (3)) plus importante que la face distale, elle-même nettement
plus convexe que la face mésiale, indiquent quil sagit très
probablement dune M3. Le protoconide est plus grand que lhypoconide et que lhypoconulide
mais sa surface est plus petite que celle de lentoconide (figure
3). Le métaconide est mésio-distalement légèrement plus court
que le protoconide mais il est bucco-lingualement légèrement plus
grand. Un important sillon mésio-distal sépare les cuspides buccales
des cuspides linguales. Il est localisé légèrement du côté buccal
par rapport à la médiane de la couronne. En dépit de lusure,
les sillons transversaux entre le protoconide et lhypoconide
dune part et entre lhypoconide et lhypoconulide dautre part
sont clairement visibles. Le sillon transversal séparant le métaconide
et lentoconide est usé, excepté à ses extrémités linguale et
buccale. Sur la figure 2 (3), la fovéa mésiale est cachée par
une concrétion sableuse mais la fovéa distale est visible. Dans
larticle de Nature (référence 1, figure 2b), la concrétion a été dégagée mais la
fovéa mésiale est difficile à discerner, probablement parce quelle
a pu avoir été très usée. Un trait remarquable de cette dent est
la présence dun métaconulide très protubérant placé disto-lingualement
du métaconide (parfois appelée sixième cuspide). De telles cuspides
sont rares chez les humains, où la fréquence est de 7% pour une
M1 (dent 36 dans la terminologie des dentistes) et denviron 2% pour
une M3 (dent 38). Une sixième cuspide se rencontre aussi dans des échantillons
de molaires inférieures de chimpanzés et de gorilles. Sachant
que le protoconide, lhypoconide et lhypoconulide sont sur la
gauche de cette dent, ils indiquent que le spécimen est une molaire
inférieure gauche. Ceci signifie quune molaire inférieure gauche
a été collée par erreur sur une mandibule droite.
Nous estimons provisoirement quil y a au minimum trois individus
dans lhypodigme des hominidés de TM 266 : lun est représenté
par le crâne holotype TM 266-02-060-1 ; un second par lhémi-mandibule
TM 266-02-154-1, ses dents étant plus usées que celles de lholotype
; un troisième par la symphyse mandibulaire TM 266-02-203, qui
partage des points communs avec lhémi-mandibule. Il reste à déterminer
si le troisième fragment de symphyse présentant des alvéoles dincisives
et de canines (TM 266-01-60-2 de la référence 1) et les dents
isolées trouvées sur ce site appartiennent ou pas à ces trois
individus là.
Deux autres spécimens dhominoïdes ont été mis au jour dans le
voisinage, une hémi-mandibule édentée (TM 247-01-02), trouvée par Mahamat
Adoum le 15 juillet 2001, et une symphyse cassée en quatre fragments
(TM 292-02-01, avec à gauche la canine, p/4-m/1, et à droite p/3,
m/1-m/2) rassemblés successivement par le premier auteur et deux
de ses collaborateurs les 15, 16 et 17 janvier 2002. Ces fossiles
représentent vraisemblablement deux individus supplémentaires
de Sahelanthropus.
Implications paléontologiques.
Après la découverte dAustralopithecus bahrelghazali (Abel) le 23 janvier 1995 dans le Sahara tchadien, la mise
au jour de Sahelanthropus (Toumaï) le 19 juillet 2001 confirme lexistence dune Lake
Chad Story, contrairement à la prédiction faite par Coppens dune
East Side Story (9). Et puis, si Toumaï nest pas un hominidé
(2), sa découverte viendrait appuyer cette hypothèse de lEast
Side Story.
Les autres fossiles de mammifères trouvés dans le même niveau,
tels des anthracothères, très abondants au Tchad alors quils
ne sont pas signalés dans les riches gisements fossilifères de
même âge dAfrique orientale (10), révèlent quil y avait un provincialisme
marqué dans les faunes de la fin du Miocène dAfrique et montre
aussi que les conditions environnementales connues du bassin du
lac Tchad pouvaient avoir permis lémergence de lespèce humaine.
La présente analyse, basée sur quelques paratypes de Sahelanthropus, est seulement un élément de la discussion au sujet du genre.
Il ne modifie pas la base du débat concernant la systématique
et la paléoécologie de Sahelanthropus.
Nous remercions le Centre National dAppui à la Recherche, un
organisme tchadien dépendant du Ministère de lEnseignement Supérieur
and ses directeurs successifs, Abakar Adoum Haggar et Baba El
Hadj Mallah, le Ministère français des Affaires étrangères, qui
employait A.B., et le SCAC (Service de coopération et daction
culturelle de lambassade de France à NDjamena pour leur appui
à la recherche et luniversité de Paris X Nanterre pour avoir
permis le long détachement de A.B. en Afrique. Nous souhaitons
particulièrement remercier nos collègues et collaborateurs tchadiens,
Mahamat Adoum, Likius Andossa, Ahounta Djimdoumalbaye, Fanoné
Gongdibé et Mackaye Hassan Taïsso, pour leur ardeur au travail
et leur excellente camaraderie sur le terrain ainsi que leur bonne
volonté pour faire progresser la connaissance scientifique en
dépit des privations du désert.
1. Brunet M., Guy F., Pilbeam D., Mackaye H.T., Likius A., Ahounta
D., Beauvilain A., Blondel C., Bocherens H., Boisserie J-R.,De
Bonis L., Coppens Y., Dejax J., Denys C., Duringer P., Eisenmann
V., Fanone G., Fronty P., Geraads D., Lehmann T., Lihoreau F.,
Louchart A., Mahamat A., Merceron G., Mouchelin G., Otero O.,
Pelaez Campomanes P., Ponce De Leon M., Rage J-C., Sapanet M.,
Schuster M., Sudre J., Tassy P., Valentin X., Vignaud P., Viriot
L., Zazzo A. et Zollikofer C. (2002). A new hominid from the Upper
Miocene of Chad, Central Africa. Nature 418, 145151.
2. Wolpoff M., Senut B., Pickford M. et Hawks J. (2002). (Communications
arising): Sahelanthropus or Sahelopithecus? Nature 419, 581582.
3. Brunet M. (2002). Reply to Sahelanthropus or Sahelopithecus? Nature 419, 582.
4. Dossier de presse, juillet 2002, Université de Poitiers, 22
pages, largement diffusé à la presse et dans lequel le surnom
de Sahelanthropus tchadensis, Toumaï, était présenté.
5. Vignaud P., Duringer P., Mackaye H.T., Likius A., Blondel C.,
Boisserie J-R., de Bonis L., Eisenmann V., Etienne M.E., Geraads
D., Guy F., Lehmann T., Lihoreau F., Lopez-Martinez N., Mourer-Chauviré
C., Otero O., Rage J-C., Schuster M., Viriot L., Zazzo A. et Brunet
M. (2002). Geology and palaeontology of the Upper Miocene Toros-Menalla
hominid locality, Chad. Nature 418, 152155.
6. Site internet http://www.chez.com/paleotchad (ou http://voila.fr/Toumaye)
7. Beauvilain A. (2003).Toumaï, laventure humaine. La Table Ronde,
Paris.
8. Brunet M., Beauvilain A., Coppens Y., Heinz E., Moutaye A.H.E.
et Pilbeam D. (1996). Australopithecus bahrelghazali, une nouvelle espèce dHominidé ancien de la région de Koro Toro
(Tchad). C. R. Acad. Sci. Paris 322, 907913.
9. Coppens Y. (1994). East Side Story: the origin of humankind.
Sci. Am. (May 1994), 8895.
10. Lihoreau F. (2003). Systématique et Paléoécologie des Anthracotheriidae
(Artiodactyla: Suiformes) du Mio-Pliocène de lAncien Monde: implications
Paléobiogéographiques. Thèse, Université de Poitiers.
|
South African Journal of Science 100, September/October 2004
|
Sahelanthropus tchadensis : Les faits
Michel Brunet and MPFT
Dans un article récemment publié dans ce journal1, Beauvilain
et Le Guellec suggèrent que notre description originale de Sahelanthropus tchadensis (2) était faussée par des identifications et des associations
de spécimens inexactes. Ces allégations nont aucun fondement.
Beauvilain et Le Guellec (1) ont proposé des informations supplémentaires
sur lhypodygme et le contexte géologique de Sahelanthropus tchadensis,
le plus ancien hominidé connu, découvert à TM266 dans lerg du
Djourab au nord du Tchad par la Mission Paléoanthropologique Franco-Tchadienne
(2,3) (MPFT). Leur intention affirmée était de décrire « les évènements
autour des découvertes elles-mêmes » (1) (p. 142), bien que ceci
ait déjà été réalisé avec prolixité par Beauvilain (4). Leur article
dans le South African Journal of Science (1) prétend que : 1) Vignaud et al. (3) ont échoué à présenter lexacte nature de la stratigraphie
du site à hominidés TM266 ; 2) Brunet et al. (2) ont négligé des
spécimens dhominidés collectés ; 3) la mandibule TM266-02-154-1
décrite par Brunet et al. (2) est en fait une chimère de fossiles
dhominidés, résultant du collage erroné dune M3 gauche sur une hémi-mandibule droite ; 4) additionnées, ces erreurs
supposées affectent le Nombre Minimum dIndividus (NMI) décompté
pour ce site.
Beauvilain et Le Guellec (1) ne remettent pas en question notre
attribution de ces fossiles au clade des hominidés (2) plutôt
quà un grand singe africain, mais tentent de démontrer que les
membres de la MPFT sont parvenus à des conclusions inexactes.
Léquipe maintient ses conclusions initiales.
Questions liées à la géologie
Étant donné lexploration géologique extensive effectuée par la
MPFT dans le Djourab depuis 1994, incluant des recherches par
georadar (3, 5-10), il est clair que lensemble de la zone fossilifère
de Toros-Ménalla (TM) ne montre aucun signe de tectonique cassante
(failles) depuis 7 Ma. Mis à part les dunes modernes, le relief
est très aplani. Dans la zone de TM, tous les petits escarpements
cités par Beauvilain et Le Guellec(1) sont les conséquences de
surcreusement éolien au pied des dunes, ce qui est très courant
dans le Sahara et dans beaucoup dautres déserts de sable. A notre
avis, ces caractéristiques géomorphologiques ne constituent pas
les « anciennes failles réactivées » quils prétendent (p. 142).
La section figurée par Vignaud et al. (3) est synthétique ; elle montre les différents faciès rencontrés,
liés aux alternances de périodes humides et sèches. La section
à TM266 est exacte telle que publiée, et tous les spécimens fossiles
cités dans les références (2) et (3) proviennent de cette section
locale publiée conjointement. Il ny a aucun doute quant à cette
provenance.
Questions liées à linventaire
Les pratiques de la MPFT en matière dinventaire et de publication
des fossiles ne diffèrent pas de celles habituellement utilisées
en paléontologie. Tous les spécimens collectés, hominidés inclus,
sont enregistrés sous un numéro dinventaire composé du numéro
de site, de lannée, du numéro du spécimen et, dans le cas dun
individu connu par plusieurs fragments, dun numéro de fragment
(par exemple, le crâne surnommé 'Toumaï' a été enregistré sous
le numéro : TM266-01-060-1). Lattribution dun tel numéro dinventaire
peut se produire à plusieurs étapes du traitement des découvertes
: 1) pour la plupart dentre elles, directement sur le terrain
après une identification précise; 2) en laboratoire, (à NDjaména
ou à Poitiers) pour tous les spécimens découverts après tamisage
ou préparation, lorsque ces spécimens sont totalement inclus dans
leur matrice dorigine. Beauvilain, géographe chargé de la logistique
au sein de la MPFT, nétait pas familiarisé avec ce processus
ni avec la façon dinclure les spécimens dans lhypodygme publié
de S. tchadensis (2).
Le premier papier décrivant le nouveau taxon S. tchadensis (2) incluait uniquement les spécimens qui ont été définitivement
identifiés en tant quhominidés par leurs caractères anatomiques.
Ces spécimens appartiennent à plusieurs individus, comme nous
lavons mentionné dans Nature (2) : le crâne holotype est lun de ces spécimens. Beauvilain
et Le Guellec (1) se demandent pourquoi une incisive très usée
(TM266-01-460) et une mandibule fragmentaire endommagée (TM266-02-203)
nont pas été incluses dans la série paratype. Elles ne lont
pas été parce que leurs affinités exactes restent à déterminer.
Au cours de leur étude, dautres restes fragmentaires ont été
attribués à des individus supplémentaires et le NMI saccroîtra
au fur et à mesure des futures fouilles et travaux de préparation.
Questions liées à la restauration et linterprétation des spécimens
Dans le désert du Djourab, la découverte des fossiles se fait
grâce à lérosion intense des sédiments dans lesquels ils sont
enfouis par laction du sable soufflé par le vent à la surface
des unités sédimentaires. Aussitôt que les fossiles sont dégagés,
la même érosion éolienne les affectent. Ils subissent les dommages
suivants : abrasion, fissures, fractures, dispersion des différents
fragments, et finalement destruction totale des spécimens sils
ne sont pas collectés presque immédiatement après le début de
leur exposition. Par exemple, le crâne TM266-01-060-1 de Sahelanthropus tchadensis était partiellement dégagé à sa découverte et a souffert dune
telle érosion liée au vent de sable. Au cours de son exposition,
il a perdu la plupart de sa denture frontale. Ainsi que Beauvilain
et Le Guellec lont rapporté (1), la canine brisée appartenant
à ce crâne a été découverte séparemment. Il ny a pas de doutes
que cette canine appartiennent à ce crâne, car, comme noté avec
exactitude par Beauvilain et Le Guellec (1) (p. 143): La dent
était composée de deux fragments qui sajustaient parfaitement
sur la racine de la canine droite . Par contre, ils ont décrit
cette canine comme étant complète, alors quil sagit en faite
de la moitié distale de la canine. Cette dent a été publiée dans
sa position naturelle, cest-à-dire remontée sur le crâne (2).
Un cas similaire sest produit pour la mandibule incriminée. La
troisième molaire droite a été séparée de la rangée dentaire par
lérosion et transportée par le vent à lendroit où elle a été
découverte, à quelques décimètres de la mandibule conservant les
autres dents. Après avoir été trouvée, nous avons établi quelle
appartenait à cette mandibule, et nous lavons recollée dans sa
position originale, en tant que M/3 droite de lhémi-mandibule
TM266-02-154-1. Toutefois, Beauvilain et Le Guellec prétendent
que cette dent a été attribuée et montée du mauvais côté.
Lanatomie occlusale de cette dent, modérément usée, est tout
à fait visible et indique sans ambiguïté aucune une troisième
molaire. Lidentification de sa latéralité a été basée sur deux
critères indépendants et suffisants, lun physique et lautre
biologique. Premièrement, il y a une concordance très claire entre
la surface inférieure de la dent et les racines conservées dans
la mandibule. Il ny a aucun doute quant à lintégrité de cette
connexion (Fig. 1A, B, C, F, G). Ceci est de plus confirmé par
la correspondance de la facette de contact préservée sur la surface
mésiale de la dent et celle de la seconde molaire restée en position
sur la mandibule. Deuxièmement, lanatomie de la troisième molaire
permet de déterminer sa latéralité sans erreur possible. Comme
pour toutes les dents dhominoïde, les cuspides vestibulaires
sont les plus usées, montrant une large cuspide plus usée (le
protoconide) marquée par la plus forte occlusion et placée mésio-vestibulairement
(Fig. 1D, E). Les bords occlusaux des cuspides linguales sont
légèrement mais clairement plus hauts, en raison de lusure plus
faible causée par les mécanismes de mastication bien connus chez
les humains actuels, les hominidés fossiles, et les grand singes
fossiles et actuels.
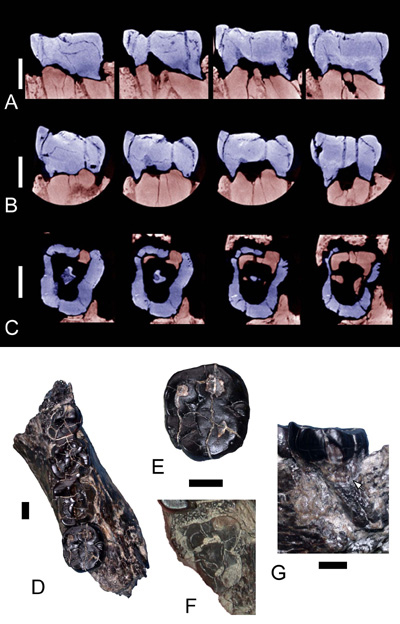
Fig. 1. Hémi-mandibule droite TM266-02-154-1 de Sahelanthropus tchadensis. A, B, and C: CT scans (the University Museum, Université de
Tokyo, Japon) au niveau de la M3. Le corps mandibulaire et les racines en place de la M3 sont en rouge clair. La couronne de la troisième molaire trouvée
séparément et prétendument du côté gauche est en bleu. Une correspondance
exacte entre la couronne de la M3 et les racines correspondantes dans lhémi-mandibule est observée.
Lespace interstitiel entre la M3 et ses racines correspond à lépaisseur de la colle utilisée
pour fixer la dent sur ses racines.
A: sections sagittales, face mésiale à droite de droite à gauche,
les CT scans ont été respectivement réalisés à 3,33 mm, 4,41 mm,
7,83 mm, et 8,70 mm du bord vestibulaire de la dent;
B, sections transversales, face linguale à droite de droite à
gauche, les CT scans ont été respectivement réalisés à 2,67 mm,
3,69 mm, 4,.11 mm, et 9,36 mm du bord mésial de la dent ;
C, sections parallèles à la surface occlusale, au niveau du collet
et en dessous, face mésiale vers le haut de droite à gauche,
les CT scans ont été respectivement réalisés à 6,93 mm, 7,14 mm,
7,44 mm, et 7,80 mm de la face occlusale de la dent. Mésialement,
les racines mésio-vestibulaire et mésio-linguale restées dans
le corps mandibulaire (voir F) sont en rouge clair et montre une
correspondance parfaite avec la couronne de la m/3 (en bleu).
D: vue occlusale du spécimen complet avec sa M3.
E: vue occlusale de la M3.
F: vue occlusale des racines de la M3.
G: vue disto-linguale du contact (flèche blanche) entre la M3 et sa racine distale.
Toutes les barres déchelle représentent 0,5 mm.
Conclusion
La contribution logistique de Beauvilain aux travaux de terrain
au Tchad est considérée avec gratitude, mais les affirmations
et les allégations quil a formulées avec Le Guellec (1) nont
aucune portée concernant linterprétation de la géologie de TM266
ou de lhypodygme, de la taxonomie ou de la position phylogénétique
de Sahelanthropus tchadensis.
Remerciements
Nous remercions les autorités tchadiennes (Ministère de l'Education
Nationale de l'Enseignement Supérieur et de la Recherche, Université
de N'djaména, CNAR). Notre gratitude va également aux ministères
français suivants pour leur soutien : Ministère français de l'Education
Nationale (Faculté des Sciences, Université de Poitiers), Ministère
de la Recherche (CNRS : SDV & ECLIPSE), Ministère des Affaires
Etrangères (DCSUR, Paris and SCAC N'djamena) ; à la Région Poitou-Charentes
; au RHOI (co-direction F.C. Howell et T.D. White) financé par
la NSF ; ainsi quà larmée française, MAM et Epervier, pour son
soutien logistique. Nous remercions particulièrement : G. Suwa
(University Museum, Université de Tokyo) pour les tomographies
scanners, C. O. Lovejoy et T. D. White pour leur discussions stimulantes,
ainsi que tous les autres membres de la MPFT ayant contribué aux
missions de terrain, et Ghislaine Florent, Carine Noël, Sabine
Riffaut pour leur aide sur le plan administratif et technique.
Références
1. Beauvilain A. and Le Guellec Y. (2004). Further details concerning
fossils attributed to Sahelanthropus tchadensis (Toumaï). South African Journal of Science 100, 142144.
2. Brunet M., Guy F., Pilbeam D., Mackaye H.T., Likius A., Ahounta
D., Beauvilain A., Blondel C., Bocherens H., Boisserie J.-R.,
Bonis L. de, Coppens Y., Dejax J., Denys C., Duringer P., Eisenmann
V., Fanoné G., Fronty P., Geraads D., Lehmann T., Lihoreau F.,
Louchart A., Mahamat A., Merceron G., Mouchelin G., Otero O.,
Pelaez Campomanes P., Ponce De Leon M., Rage J.-C., Sapanet M.,
Schuster M., Sudre J., Tassy P., Valentin X., Vignaud P., Viriot
L., Zazzo A. and Zollikofer C. (2002). A new hominid from the
Upper Miocene of Chad, Central Africa. Nature 418, 145151.
3. Vignaud P., Duringer P., Mackaye H.T., Likius A., Blondel C.,
Boisserie J.-R., Bonis L. de, Eisenmann V., Etienne M.-E., Geraads
D., Guy F., Lehmann T., Lihoreau F., Lopez-Martinez N., Mourer-Chauviré
C., Otero O., Rage J.-C., Schuster M., Viriot L., Zazzo A., Brunet
M. (2002). Geology and palaeontology of the Upper Miocene Toros-Menalla
hominid locality, Chad. Nature 418, 152155.
4. Beauvilain A. (2003). Toumaï, laventure humaine. La Table
Ronde, Paris.
5. Brunet M., Beauvilain A., Coppens Y., Heintz E., Moutaye A.H.E.
and Pilbeam D. (1995). The first Australopithecine 2 500 kilometres
west of the Rift Valley (Chad). Nature 378, 273274.
6. Brunet M., Beauvilain A., Geraads D., Guy F., Kasser M., Mackaye
H.T., Maclatchy L. M., Mouchelin G., Sudre J. and Vignaud P.,
1998. Tchad: découverte d'une faune de Mammifères du pliocène
inférieur. Comptes Rendus de l'Académie des Sciences Paris 326, 153158.
7. Brunet, M. and M.P.F.T. (2000). Chad: discovery of a Vertebrate
fauna close to the Mio-Pliocene boundary. Journal of Vertebrate Paleontology 20 (1), 205209.
8. Schuster M. (2002). Sédimentologie et paléoécologie des séries
à vertébrés du paléolac Tchad depuis le Miocène supérieur. Ph.D.
thesis, Université Louis Pasteur, Strasbourg.
9. Ghienne J.M., Schuster M., Bernard A., Duringer Ph. and Brunet
M. (2001). The Holocene giant Lake Chad revealed by digital elevation
models. Quaternary International 87, 8185
10. Schuster M., Duringer Ph., Ghienne J.F., Beauvilain A., Mackaye
H.T., Vignaud P. and Brunet M. (2003). Discovery of coastal conglomerates
around the Hadjer El Khamis Inselbergs (Western Chad, Central
Africa) : a new evidence for lake Mega-Chad episods. The Costal
conglomerates of Lake Mega-Chad. Earth Surfaces Processes and Landforms 28 (10), 10591069.
LETTRE INTERNATIONALE DE SOUTIEN
1er juillet 2004
Dr. Graham Baker
Editor South African Journal of Science
Cher Graham,
Un article récemment publié dans le South African Journal of Science a mis en doute lidentité dun fossile tchadien exceptionnel
ainsi que la compétence de léquipe qui la décrit dans Nature.
En pièce jointe, vous trouverez une note quun groupe de paléoanthropologues
professionnels et nous-même voudrions publier dans votre journal
pour clarifier lidentification du spécimen concerné. Tous nos
co-auteurs ont reçu larticle de Beauvilain et Le Guellec paru
dans le SAJS en format pdf, ainsi que des images scanners en format
jpeg et des photographies précises de la dent concernée (artificiellement
isolée de la mandibule, de façon à ce que sa position soit déterminée
exclusivement à partir de la morphologie de la couronne dentaire).
Le-mail que nous avons envoyé à nos collègues le 22 juin dernier
est reproduit ci-dessous.
Comme vous pouvez le constater, à ce jour, 27 auteurs de 13 pays
différents ont répondu, tous identifiant sans équivoque la dent
comme étant une droite. Beaucoup de ces co-auteurs ont fourni
des arguments anatomiques pour justifier leur identification,
et plusieurs ont exprimé leur surprise sur le fait quun tel article
ait pu passer à travers le processus dévaluation sans avoir été
mis en cause.
Nous savons que le Professeur Brunet fournit une réponse indépendante
à Beauvilain et Le Guellec, et nous espérons que notre contribution
pourra être publiée aux côtés de celle-ci et des excuses que le
Dr. Martin Pickford vous a envoyées par e-mail.
Cordialement,
F. Clark Howell Tim D. White
Cher Monsieur,
Nous, soussignés, avons examiné en détail les photographies et
images digitales de la couronne dune troisième molaire fossilisée
provenant du Miocène supérieur du Tchad. Cette dent a été originellement
identifiée par ses découvreurs (Brunet et al., 2002) comme une troisième molaire inférieure droite. Un article
récent écrit par Beauvilain et Le Guellec (2004) prétendait que
cette dent avait été mal identifiée et quelle était en fait une
troisième molaire inférieure gauche. Sur la base de la morphologie
de la couronne visible sur les images que nous avons examinées,
nous confirmons que cette dent est bien une molaire droite, ainsi
que lont publié Brunet et al. (2002).
Signé : F. Clark Howell
Laboratory for Human Evolutionary Studies,
Museum of Vertebrate Zoology,
The University of California at Berkeley,
Berkeley, CA 94720, U.S.A.
E-mail: fchlhes@socrates.berkeley.edu
Tim D. White
Laboratory for Human Evolutionary Studies,
Museum of Vertebrate Zoology and
Department of Integrative Biology,
The University of California at Berkeley,
Berkeley, CA 94720, U.S.A.
E-mail: timwhite@socrates.berkeley.edu
David R. Begun
Department of Anthropology,
University of Toronto,
Toronto, ON M5S 3G3,Canada.
E-mail: begun@chass.utoronto.ca
Yaowalak Chaimanee
Paleontology Section,
Bureau of Geological Survey,
Department of Mineral Resources,
Rama VI Road, Bangkok 10400, Thailand.
E-mail: yaowalak@dmr.go.th
Marie-Antoinette de Lumley
Institut de Paléontologie Humaine.
75013 Paris, France.
E-mail: prehist@mnhn.fr
Philip D. Gingerich
Museum of Paleontology,
University of Michigan,
Ann Arbor, MI 48109-1079, U.S.A.
E-mail: gingeric@umich.edu
Colin Groves
School of Archaeology & Anthropology,
Australian National University,
Canberra, A.C.T. 0200, Australia.
E-mail: Colin.Groves@anu.edu.au
Erksin Güleç
Department of Physical Anthropology and Paleoanthropology,
University of Ankara,
06100, Sihhiye, Ankara, Turkey.
E-mail: Erksin.Gulec@humanity.ankara.edu.tr
Yohannes Haile-Selassie
Cleveland Museum of Natural History,
1 Wade Oval Drive, Cleveland, OH 44106, U.S.A.
E-mail: yhailese@cmnh.org
Leslea Hlusko
Department of Integrative Biology,
The University of California at Berkeley,
Berkeley, CA 94720, U.S.A.
E-mail: hlusko@socrates.berkeley.edu
Jean-Jacques Jaeger
Equipe Phylogénie, Paléobiologie & Paléontologie
I.S.E.M.,
CNRS-Universite Montpellier II, UMR 5554,
Cc 064 Place Eugene Bataillon,,
34095 MONTPELLIER CEDEX 5, France.
E-mail: jaeger@isem.univ-montp2.fr
Jay Kelley
Department of Oral Biology,
College of Dentistry,
University of Illinois at Chicago,
Chicago, IL 60612, U.S.A.
E-mail: jkelley@uic.edu
Meike Köhler
Institute de Paleontologia M. Crusafont,
c/ Escola Industrial 23,
08201 Sabadell, Barcelona, Spain.
E-mail: kohlerlm@diba.es
Wu Liu
Institute of Vertebrate Paleontology and Paleoanthropology,
Chinese Academy of Sciences,
Beijing 100044, China.
E-mail: liuwu@ivpp.ac.cn
David Lordkipanidze
Georgian State Museum,
0105 Tbilisi, Georgia.
E-mail: geonathist@ip.osgf.ge
C. Owen Lovejoy
Matthew Ferrini Institute for Human Evolutionary Research,
Department of Anthropology,
Kent State University,
Kent, OH 44242, U.S.A.
E-mail: OLovejoy@aol.com
Lawrence B. Martin
Department of Anthropology and of Anatomical Sciences,
The Graduate School,
Suite 2401, Computer Science Building,
Stony Brook University, NY 11794-4433, U.S.A.
E-mail: Lawrence.Martin@stonybrook.edu
Monte L. McCrossin
Department of Sociology and Anthropology,
MSC 3BV,
New Mexico State University,
P.O. Box 30001, Las Cruces, NM 88003-8001, U.S.A.
E-mail: mmccross@nmsu.edu
Salvador Moyà-Solà
Institute de Paleontologia M. Crusafont,
c/ Escola Industrial 23,
08201 Sabadell, Barcelona, Spain.
E-mail:moyass@diba.es
Lorenzo Rook
Dipartimento di Scienze della Terra,
Università di Firenze,
via G. La Pira, 4, 50121 Firenze, Italy
E-mail: Lrook@geo.unifi.it
Pat Smith
Department of Anatomy,
The Hebrew University Hadassah Medical School,
Jerusalem, Israel.
E-mail: pat@cc.huji.ac.il
Gen Suwa
The University Museum
The University of Tokyo
Hongo, Bunkyo-ku
Tokyo, 113-0033, Japan
E-mail: suwa@um.u-tokyo.ac.jp
Mark Teaford
Center for Functional Anatomy and Evolution,
Johns Hopkins University School of Medicine,
Baltimore, MD 21205, U.S.A.
E-mail: mteaford@jhmi.edu
Phillip V. Tobias
School of Anatomical Sciences,
University of the Witwatersrand,
Johannesburg, South Africa.
E-mail: tobiaspv@anatomy.wits.ac.za
Alan Walker
Departments of Anthropology and Biology,
The Pennsylvania State University,
University Park, PA 16802, U.S.A.
E-mail: axw8@psu.edu
Phil Walker
Department of Anthropology,
The University of California at Santa Barbara,
Santa Barbara, CA 93106, U.S.A.
E-mail: pwalker@anth.ucsb.edu
Steven Ward
Department of Anatomy
NEOUCOM
Rootstown, OH 44272-0095
E-mail: scw@neoucom.edu
Réponse de Beauvilain et Le Guellec
Brunet et al (1) critiquent trois points importants de notre article
(2) : a) lorigine tectonique de lescarpement de Toros-Menalla,
b) la détermination du côté dune molaire isolée qui a été collée
sur une mandibule droite trouvée sur le site de TM 266 et attribuée
à Sahelanthropus tchadensis et c) linventaire des fossiles. Nous répondons à chacune de
ces critiques et concluons que nos hypothèse et interprétation
originelles sont bien fondées.
Les problèmes géologiques dans le Djourab.
Brunet et al (1) rendent compte quun géoradar na rapporté aucune
évidence de failles dans la région de Toros-Menalla et ils attribuent
la formation de lescarpement à un surcreusement par le vent.
Bien queffectivement la déflation éolienne cause des dépressions
dans les déserts, de telles cuvettes sont rarement linéaires,
étant habituellement ondulées dans leurs contours ou en forme
de cuiller. Le fait que lescarpement en question soit rectiligne
sur une distance de plus de 40 km suggère une origine tectonique.
Ailleurs dans la cuvette tchadienne (3) une activité néotectonique
est évidente comme le rivage occidental du lac Tchad qui est rectiligne
NNW-SSE sur environ 250 km, direction que prolonge la vallée de
la Dilia au Niger sur une distance encore plus grande. La vallée
du Bahr el Ghazal est également rectiligne NNE-SSW sur plus de
450 km avec dans sa partie aval une falaise sur sa rive gauche.
Cette activité néotectonique réactive fréquemment danciennes
fractures (4).
Un géoradar a été effectivement utilisé en janvier 1999 mais seulement
sur des sites situés à lEst du Bahr el Ghazal (la région de Toros-Menalla
est à lOuest). Sur des dunes fossiles ou récentes, il a révélé
des réflectances sur seulement un ou deux mètres dépaisseur (5)
et dans des grès il « na pu donner une image à plus dun mètre
de profondeur » (5). Ce géoradar nest pas conçu pour repérer
les témoignages de failles et il nétait pas utilisé pour chercher
des failles dans le Djourab. Cet échec du matériel pour découvrir
des failles dans cette région ne fournit pas un argument solide
contre notre opinion que les structures linéaires qui se rencontrent
dans cette région sont vraisemblablement dorigine tectonique.
Par conséquent nous maintenons notre opinion initiale que lescarpement
de Toros-Menalla, qui a donné tous les fossiles de Sahelanthropus tchadensis, doit ses origines à lactivité tectonique. Il a été par la suite
érodé par le vent mais celui-ci a seulement modifié sa forme plutôt
quil en soit la cause originelle.
Restauration et interprétation des fossiles de Sahelanthropus.
Les fossiles du Djourab sont bien minéralisés mais sont souvent
abrasés par le vent de sable. De forts écarts quotidiens de température
existent, qui peuvent varier de 50° C pour la température de lair
au niveau du sol en milieu de journée à des gelées blanches la
nuit. A midi, les cailloux et les fossiles à la surface du sol
peuvent être trop chauds pour être touchés. Ces écarts aboutissent
au fissurage des fossiles qui ont habituellement des surfaces
de fracture planes plutôt que fortement courbées. Les croûtes
qui recouvrent les fossiles se présentent soit sous forme dune
matrice siliceuse grise, dure, très adhérante au fossile, inattaquable
par voie chimique, soit sous forme dune concrétion ferrugineuse,
colorée par des sels de manganèse et souvent détachée du fossile
lui-même par un petit espace occupé par du sable (photo 1).
Le crâne de Toumaï, qui a été trouvé à lenvers sur le sable,
a été protégé de lérosion par ce deuxième type de croûte tandis
que la demi-mandibule droite TM 266-02-154-1 portait trace de
la matrice grise (photo 2) qui enrobait également la base de la
M3. Aussi nous considérons que le crâne dune part, la mandibule
et la M3 dautre part nont pas été fossilisés dans les mêmes dépôts et
par conséquent ne sont probablement pas contemporains.
La reconstitution la plus simple du passé de la M3 et de la demi-mandibule droite est leur fracture avant leur enfouissement
et leur fossilisation voici 6 à 7 millions dannées comme le montre
la croûte couvrant la surface cassée de la racine distale de la
M3 (photo 2). Ultérieurement, elles sont encroûtées près de la surface.
Récemment exhumées par le vent, labrasion a dégagé en grande
partie leur matrice et a poli leur surface tandis que les écarts
thermiques ont fait éclater certaines dents. La M3 a été probablement le dernier specimen à être érodé hors des
sédiments car sa surface est bien conservée. Inversement, lautre
demi-mandibule droite (TM 266-02-203), découverte en mars 2002
sans dent à une douzaine de mètres de la mandibule et de la M3, a été la première à être exposée à lair.
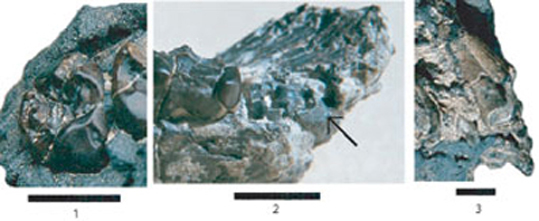
Fig. 1-3. Photographies de spécimens attribués à Sahelanthropus tchadensis (prises le 19 juillet 2001 à 8 h. pour le crâne de Toumaï) :
1) la M3 du crâne ;
2) vue linguale oblique des racines de la M3 de la mandibule (noter en particulier la surface fracturée relativement
plane de la racine distale qui se courbe distalement et buccalement
à un niveau constant (flèche noire) et comparez-la avec la surface
convexe antério-postérieure indiquée par une flèche sur limage
fournie par Brunet et al (1) ;
3) racine de la canine droite du maxillaire de Toumaï. (échelle
: 10 mm)
Considérations biologiques concernant la M3.
Lun des arguments présentés par Brunet et al (1) concernant la
détemination du côté de la M3 isolée est le mode dusure. Cependant, lexamen de la dernière
molaire droite du crâne de Toumaï (photo 1) indique que nous devons
être prudents sur linterprétation, en général, de lusure des
pointes cuspidiennes. Curieusement, dans ce cas particulier, lusure
des cuspides vestibulaires est similaire de celle des cuspides
linguales.
Cest pourquoi, dès le début de cette étude (2), nous avons accordé
davantage dimportance aux sillons, qui sont moins affectés par
lusure durant la vie (et plus récemment par lérosion éolienne)
et sont par conséquent mieux préservés que les pointes cuspidiennes.
Premièrement, la ressemblance entre la M3 et une molaire humaine « moderne » est frappante. Lalignement
sur une courbe régulière des trois cuspides vestibulaires (protoconide,
hypoconide, hypoconulide) et leur taille décroissante dans le
sens mésio-distal constituent un critère déterminant. Deuxièmement,
la colonne inter-cuspidienne, que nous avons appelée métaconulide
dans notre article, évoque plus une variation morphologique de
la face linguale que de la face vestibulaire. Troisièmement, lorientation
du sillon principal disto-vestibulaire correspond bien à la dynamique
articulaire occlusale de la diduction non travaillante des anthropomorphes,
véritable réplique en creux de léminence occlusale de la crête
oblique maxillaire ou « pont démail » (6).
Une seconde série darguments employés par Brunet et al (1) pour prouver que la M3 sadapte sur la droite de la mandibule est une série de vues
scannées dans laquelle la surface brisée de la base de la dent
et les racines dans la mandibule seraient compatibles, étant seulement
séparées par une couche de colle fine et continue. Il faut reconnaître
le travail minutieux qui a permis ladaptation avec précision
de la couronne de la M3 sur les racines de lhémi-mandibule. La dure gangue sableuse
qui couvrait la base de la molaire isolée quand elle a été découverte
a dabord dû être dégagée puis lespace maintenant occupé par
la colle a dû être apprêté millimètre par millimètre. Un travail
similaire a été requis pour les parties de la racine dans la mandibule
qui étaient aussi encroûtées (photo 2). Dune façon générale,
une coupe au collet de deux molaires mandibulaires symétriques
donne des surfaces radiculaires aux formes superposables à un
millimètre près. Il nest donc pas surprenant, dans ce cas précis,
quune dent de gauche ait pu sembler correspondre à des racines
de la partie droite de la mandibule.
La canine droite supérieure de Toumaï
Nous présentons nos excuses pour la mauvaise interprétation possible
de notre phrase concernant la canine droite de Toumaï dont la
racine fait bien partie intégrante du crâne comme le montre la
Fig. 3. Par contre, nous navons fait que rapporter par écrit
que la couronne trouvée en novembre 2001 sadaptait parfaitement
sur la racine, étant nous-même dans le Djourab et le crâne à lUniversité
de Poitiers.
Les problèmes de conservation.
Inscrire les numéros du catalogage sur les fossiles est une activité
quotidienne sur le terrain. Dans le cas dun site aussi important
que TM 266, tous les fossiles sont collectés. Cest pourquoi,
de juillet à décembre 2001, 52 pièces post-crâniennes, dont le
groupe zoologique navait pu être déterminé sur le terrain ont
été cataloguées en espérant que quelques unes dentre elles pourraient
appartenir à des Sahelanthropus. Parmi ces pièces, 36 concernent des os longs (tibia, fémur, humérus
et cubitus) sous forme soit de spécimens intacts, soit de diaphyses
cassées. Considérant lexcellent état de conservation du crâne
de Toumaï, un examen approfondi de ces os pourrait apporter dintéressantes
informations car nous estimons probable que des fossiles post-crâniens
dun grand primate soient présents sur le site, quoique rien nait
été rapporté jusquà présent à ce sujet. Notons que dans linventaire remis à NDjamena par lUniversité
de Poitiers en décembre 2001, un seul fossile a été ajouté au
catalogue établi sur le terrain. Il concerne le spécimen TM 266-01-447
(une M3 droite selon Brunet et al (7)) alors que le catalogue mentionne « Classification : primate
; Description : fragments morceaux racines M1/M3 ; Dépôt : Poitiers
(reliquat tamis) ». Ces spécimens sont restitués au CNAR à NDjamena
le 30 janvier 2002.
Conclusion
Nous ne voyons aucune raison irréfutable de modifier radicalement
notre hypothèse au sujet de la géomorphologie de la région de
Toros-Menalla, ni notre interprétation de la M3 gauche isolée et de lhémi-mandibule droite endommagée provenant
du site TM 266 et attribuées à Sahelanthropus tchadensis. Enfin, étant donné lexcellente préservation des fossiles du
site, nous considérons comme vraisemblable que la collection déléments
post-crâniens qui ont été collectés puisse bien contenir quelques
spécimens appartenant à Sahelanthropus.
(1) Brunet et MPFT (2004). Sahelanthropus tchadensis : the facts. South African Journal of Science, 100,
(2) Beauvilain A., Le Guellec Y. (2004). Further details concerning
fossils attributed to Sahelanthropus tchadensis (Toumaï). South African Journal of Science, 100, 142-144.
(3) Morin S. (2000). Géomorphologie. In Atlas de la province Extrême-Nord
Cameroun, pp. 7-17, IRD-LCA/France MINREST-INC :Cameroun.
(4) Neev D., Hall J.K., Saul J.M. (1982). The Pelusium Megashear
System Across Africa and associated Lineament Swarms. Journal of Geophysical Research, 87, B2,1015-1030.
(5) Schuster M. (2002). Sédimentologie et paléoécologie des séries
à vertébrés du paléolac Tchad depuis le Miocène supérieur. Thèse,
Université de Strasbourg.
(6) Alain Lautrou, Université de Paris V René Descartes, auteur
de Anatomie dentaire, Masson, Paris.
(7) Brunet et al. (2002). Nature 418, 145151.
|
South African Journal of Science 101, March/April 2005
|
Nouvelle correspondance concernant Sahelanthropus tchadensis
Alain Beauvilain and Yves Le Guellec
Monsieur - Les lecteurs suivant le débat au sujet de Sahelanthropus tchadensis dans cette revue scientifique doivent savoir que largument principal
de la réplique de M. Brunet et al. à notre article est fondé sur une série de CT scans et de photographies
numériques qui révèlent des contradictions entre la figure et
sa légende (1). Dans notre réplique à cette réponse (2), nous
navons pas mentionné ces contradictions parce que nous avons
considéré quil était peu probable que ces documents soient publiés
tel que soumis. Cependant, il est clair que :
1) Il y a une erreur dans les échelles puisque des dents dhominidé
de quelques millimètres de long, ainsi quil est proposé par la
légende, sont inconnues. Lerreur est de 1 à 10.
2) Il y a une erreur dans lorientation des CT scans A dont la
légende est : « sections sagittales, face mésiale à droite
».
La racine de la M3, qui sincline vers la droite, prouve que ce nest pas la face
mésiale qui est à droite mais la face distale. En effet, pour
la troisième molaire, la racine mésiale est verticale et la racine
distale est inclinée vers larrière de la mandibule où ne se trouve
plus de dent pour entraver son enracinement. Ce point de vue est
parfaitement confirmé par la photographie G figurant sur cette
même planche. De plus, sur les deux images centrales, la facette
interstitielle plate, caractéristique de la surface mésiale, se
trouve bien à gauche, et la surface interstitielle bombée, caractéristique
de la surface distale, à droite.
Lexamen de la section sagittale « réalisée à 3,33 mm du bord
vestibulaire de la dent » (la coupe la plus à droite des CT scans)
et des photographies E et G, et particulièrement les lignes de
fracture et les marques dusure quelles présentent, montre que
les sections sagittales ont été en réalité réalisées à partir
du bord lingual de la dent.
3) une erreur de positionnement latéral des CT scans B dont la
légende « sections transversales, face linguale à droite de
droite à gauche, les CT scans ont été respectivement réalisés
à 2,67 mm, 3,69 mm, 4,11 mm, et 9,36 mm du bord mésial de la dent
».
Si la face linguale est à droite, la photographie présente une
dent de gauche. Auquel cas son commentaire devient incompréhensible
par rapport aux CT scans. Par ailleurs, les mécanismes de mastication
des hominoïdes admettent des exceptions. Linclinaison du plan
occlusal est dépendante de la préparation des vestiges de racines
encore présentes sur la face cervicale de la couronne et des racines
solidaires du corps mandibulaire, préalablement au collage. T.D.
White expose bien les difficultés de tout collage (2). Dune manière
générale, retrouver les contacts parfaits entre la couronne et
les racines dune dent, indépendantes depuis des millions dannées,
encroûtées séparément dans un ciment très dur avant dêtre soumise
récemment à une intense abrasion éolienne et à des écarts de température
qui ont fait éclater de nombreux morceaux de dents, demeure très
hypothétique.
La comparaison avec la photographie E montre que, de droite à
gauche, ces sections transversales ont été en fait réalisées à
partir du bord distal de la dent.
4) une erreur de positionnement latéral des CT scans C dont la
légende est « sections parallèles à la surface occlusale, au niveau
du collet et en dessous, face mésiale vers le haut de droite
à gauche, les CT scans ont été respectivement réalisés à 6,93
mm, 7,14 mm, 7,44 mm, et 7,80 mm de la face occlusale de la dent.
Mésialement, les racines mésio-vestibulaire et mésio-linguale
restées dans le corps mandibulaire sont en rouge clair et montre
une correspondance parfaite avec la couronne de la M3 (en bleu) ».
Selon la légende, la photo de droite est donc réalisée le plus
près de la surface occlusale de la dent et la photo de gauche
réalisée le plus près du corps mandibulaire. Or les racines et
le corps mandibulaire apparaissent bien sur la photographie de
droite, la plus proche de la face occlusale de la dent, et disparaissent
pratiquement de la photographie de gauche indiquée comme ayant
été prise au plus près du corps mandibulaire.
Regrettons quaucun scan, ni aucune photographie ne montre la
face latérale de la M3 et ce petit cuspide que nous avons appelé métaconulide (1 sur
les photographies), élément particulièrement original de cette
dent. Incidemment, notons que sur la vue occlusale ce cuspide
accessoire semble porter une marque (traînée transversale blanche
sur limage de gauche) qui nexistait pas lors de sa découverte
(image de droite).
Par ailleurs, la présence supposée dune facette de contact entre
la M2 et la M3 ne peut être utilisée pour démontrer que les deux dents appartiennent
à un même individu pour la simple raison que les deux tiers de
la face distale de la M2, qui forme normalement cette facette, sont manquants ayant été
éclatés par les agents atmosphériques. Le tiers restant de la
surface distale a subi labrasion éolienne (figure, image de gauche).
Il nexiste donc aucune preuve du contact entre ces deux dents
(2 sur la photographie). Il nest donc pas possible daffirmer,
comme il en est fait mention dans la réponse de la MPFT, la correspondance
de la facette de contact préservée sur la surface mésiale de la
dent et celle de la seconde molaire restée en position sur la
mandibule.
La conservation des deux fossiles est très différente, M1 et M2 en place dans la mandibule ont été très dégradées par les aléas
atmosphériques alors que la M3 est pratiquement intacte. De plus, si la M3 appartient à la mandibule, le gradient dusure due à la mastication
doit diminuer de la M1 à la M3. Dans ce cas précis, la M3 apparaît comme étant la dent la plus usée, montrant de plus grandes
surfaces de dentine que la M2.
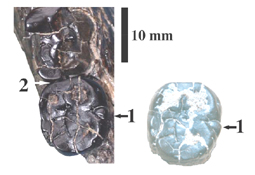
Légende de la figure : Les deux molaires, M2 et M3, après préparation (à gauche) et M3 au moment de sa découverte.
1) la petite cuspide que nous appelons métaconulide ;
2) ici napparaît pas lévidence dun contact physique entre les
deux dents.
Dans ces conditions, il est étonnant quaucun des 28 co-signataires,
regroupés sous le sigle MPFT, ne se soit aperçu des multiples
problèmes posés par leur réponse. Comment aussi comprendre que
28 autres signataires incluant de nombreux éminents paléoanthropologues,
après avoir "examiné en détail les photographies et images digitales
de la couronne dune troisième molaire fossilisée provenant du
Miocène supérieur du Tchad" puissent co-signer une lettre internationale
de soutien à la MPFT, lettre dépourvue de tout contenu scientifique
(4).
Outre que lhistoire des sciences napporte aucun précédent à
ce type de lettre, lun de ces co-signataires a récemment proposé
une liste de recommandations déontologiques (5). Pour ce cas précis,
nous ne retiendrons que les recommandations suivantes parmi un
riche échantillon de ce qui est un article essentiellement méthodologique
: "ne pas publier sans avoir dabord examiner loriginal", "ne
pas juger de la qualité dune communication par ses auteurs",
"lire de manière critique toute communication scientifique", "ne
pas rejeter les communications ou accorder des applications pour
des raisons personnelles ou politiques", ainsi que lobservation
que "
les micro et les macro-tomographies informatisées, le microscope
à balayage électronique, les coupes laser,
, les images 3D, et
lanalyse précise ne sont que des outils. Ils nont pas de qualités
magiques."
Nous souscrivons pleinement à cet excellent conseil et nous recommandons
quil soit pris au sérieux afin de réduire la possibilité dune
méprise érudite en ce domaine.
Alain Beauvilain
Université de Paris X Nanterre, 200 avenue de la République, 92000
Nanterre Cedex, France.
Yves Le Guellec
Rue du Manoir, 76190 Yvetot, France.
1. Brunet M. et al. (2004). Sahelanthropus tchadensis : the facts. S. Afr. J. Sci. 100, 443445.
2. Beauvilain A. And Le Guellec Y. (2004). S. Afr. J. Sci. 100, 445446.
3. White T.D. (2000). Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia. Am. J. Phys. Anthropol. 111, 4568.
4. Howell F.C. et al. (2004). S. Afr. J. Sci. 100, 446.
5. White T.D. (2000). A view on the science : physical anthropology
at the millenium. Am. J. Phys. Anthropol. 113, 287292.
Note de M. Brunet et al.
Nous remercions le Docteur Beauvilain pour avoir indiqué les erreurs
de la légende de notre planche illustrée mais souhaitons indiquer
que l'erreur typographique de l'échelle et l'inversion accidentelle
des informations concernant la latéralité de notre légende n'affectent
pas les données anatomiques et l'interprétation que nous avons
données pour le matéiel présenté. La légende correcte doit être
lue comme suit :
Fig. 1. Hémi-mandibule droite TM266-02-154-1 de Sahelanthropus tchadensis. A, B, and C: CT scans (the University Museum, Université de
Tokyo, Japon) au niveau de la M3. Le corps mandibulaire et les racines en place de la M3 sont en rouge clair. La couronne de la troisième molaire trouvée
séparément et prétendument du côté gauche est en bleu. Une correspondance
exacte entre la couronne de la M3 et les racines correspondantes dans lhémi-mandibule est observée.
Lespace interstitiel entre la M3 et ses racines correspond à lépaisseur de la colle utilisée
pour fixer la dent sur ses racines.
A: sections sagittales, face mésiale à gauche de gauche à droite, les CT scans ont été respectivement réalisés à 3,33 mm, 4,41
mm, 7,83 mm, et 8,70 mm du bord vestibulaire de la dent;
B : sections transversales, face linguale à gauche de gauche à droite, les CT scans ont été respectivement réalisés à 2,67 mm, 3,69
mm, 4,11 mm, et 9,36 mm du bord mésial de la dent ;
C : sections parallèles à la surface occlusale, au niveau du collet
et en dessous, face mésiale vers le haut de gauche à droite, les CT scans ont été respectivement réalisés à 6,93 mm, 7,14
mm, 7,44 mm, et 7,80 mm de la face occlusale de la dent. Mésialement,
les racines mésio-vestibulaire et mésio-linguale restées dans
le corps mandibulaire (voir F) sont en rouge clair et montre une
correspondance parfaite avec la couronne de la m/3 (en bleu).
D : vue occlusale du spécimen complet avec sa M3.
E : vue occlusale de la M3.
F : vue occlusale des racines de la M3.
G : vue disto-linguale du contact (flèche blanche) entre la m/3
et sa racine distale.
Toutes les barres déchelle représentent 0,5 cm.
|
|
